Филогения*
— часть биологии, рассматривающая вопрос о происхождении организмов друг от друга. Как особая отрасль Ф. почти нигде не преподается и не излагается, хотя входит как необходимая составная часть почти в каждый отдел научного курса. Впрочем, в Иенском университете имеется особая кафедра Ф. (Ritter-Professur f ü r Phylogenie). В настоящее время вряд ли найдется хоть один биолог, который не принимал бы учение о Ф. или принимал его лишь в ограниченном смысле, напр. в пределах отдельных типов, как это делали Кювье и Бэр (см. Трансформизм и Теория типов). Достаточно доказать, что хотя бы один вид произошел от другого, и мы, конечно, должны общий вывод, сделанный из такого даже единичного факта, распространить на все существующие виды, ибо законы, управляющие филогенетическим развитием, одинаковы для всего организованного мира. Типы есть такое же совершенно условное деление, как и всякое другое, и, конечно, реального существования им приписать нельзя. Реальны лишь различия между отдельными особями, различия, которые в минимальной их степени имеют значение индивидуального уклонения, а в максимальной — границы между типами. Для определения филогенетического родства можно руководиться различными приемами. Конечно, сравнительная анатомия и изучение переходных форм оказывает громадные услуги, но не меньшее значение имеет и сравнительная эмбриология, опирающаяся на сходство ранних стадий высших животных со взрослыми формами низших (см. Филогенетический закон). Наконец, обильный материал дает нам и палеонтология. Если выяснение филогенетических отношений далеко не закончено, то этому причиной неполнота наших знаний. Большинство переходных форм вымерло, и многие из них, вероятно, вымерли, не оставив никаких остатков. Да и современная фауна, особенно морская, далеко не изучена окончательно: достаточно сказать, что до последнего времени мы продолжаем находить как в глубине моря, так и на материках такие формы, которые или считались существующими лишь в ископаемом состоянии, или не были известны вовсе. Переходя к Ф. животного царства, отметим, что после неудачной попытки представить ее в виде восходящей лестницы (см. Теория животной лестницы) зоологи, сначала в лице Эразма Дарвина, а потом главным образом в лице Ламарка, перешли к изображению филогенетических отношений в виде разветвленного дерева (см. Трансформизм). Особую разработку этот вопрос получил в последарвиновский период, причем с наибольшей ясностью вопрос о Ф. был поставлен Геккелем. Неполнота наших знаний приводит к тому, что каждое филогенетическое дерево заключает в себе значительную долю субъективности. Конечно, мы ограничимся рассмотрением Ф. типов (см.). Относительно перехода от простейших (Protozoa) к многоклеточным (Metazoa) мы имеем несколько гипотез. Две наиболее имевшие успех гипотезы берут за исходную точку шарообразную (похожую на Volvox или Magosphaera) колонию, но одна гипотеза (Геккеля, Grastraea-theorie) принимает, что внутренний пласт многоклеточных образовался через впячивание или вворачивание одной части сферы в другую (см. Гаструлация), а другая гипотеза (Раrеnchymella-theorie Мечникова) предполагает, что этот пласт образовался через внедрение отдельных клеток внутрь сферы (иммиграция).
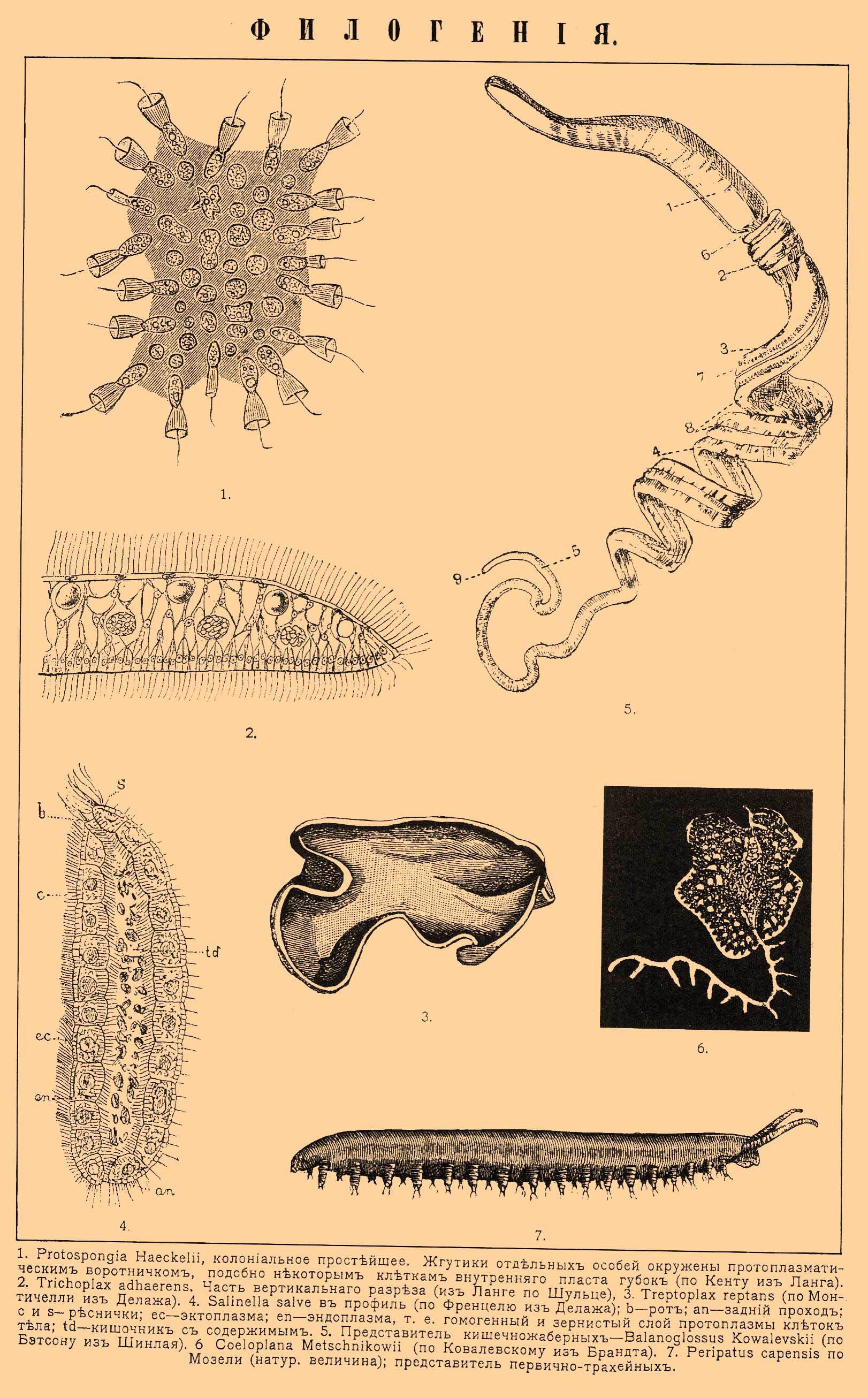
ФИЛОГЕНИЯ. 1. Protospongia Haeckelii, колониальное простейшее. Жгутики отдельных особей окружены протоплазматическим воротничком, подобно некоторым клеткам внутреннего пласта губок (по Кенту из Ланга). 2. Trichoplax adhaerens. Часть вертикального разреза (из Ланге по Шульце). 3. Treptoplax reptans (по Монтичелли из Делажа). 4. Salinella salve в профиль (по Френцелю из Делажа); b — рот; аn — задний проход; c и s — реснички; ес — эктоплазма; еn — эндоплазма, т. е. гомогенный и зернистый слой протоплазмы клеток тела; td — кишечник с содержимым. 5. Представитель кишечножаберных Balanoglossus Kowalevskii (по Бэтсону из Шинлая). 6. Coeloplana Metschnikowii (по Ковалевскому из Брандта). 7. Peripatus capensis, по Мозели (натур. величина); представитель первичнотрахейных.
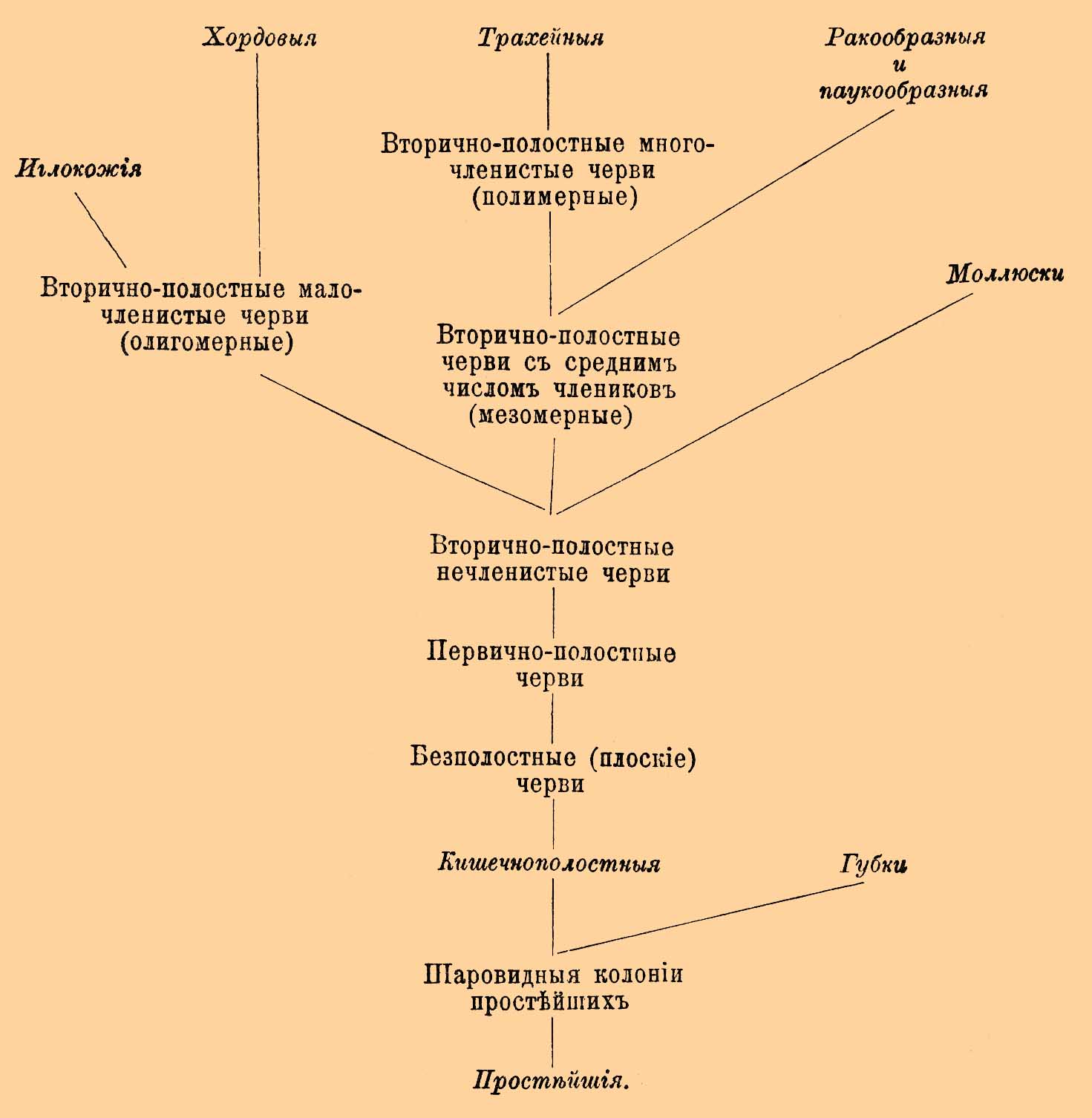
Обе гипотезы могли бы опереться на некоторые переходные формы, для которых ван Бенеден предложил название Mesozoa, но вряд ли только все формы, описываемые под этим общим наименованием, представляют естественную группу. Многие из них произошли вследствие упрощения под влиянием паразитического образа жизни из более сложных форм, другие, может быть, представляют личиночные стадии. К числу последних относятся, вероятно, описанный Монтичелли (Monticelli, 1896) организм под именем Pemmatodiscus, представляющий типичную гаструлу и живущий в ткани диска одной медузы (Rhizostoma pulmo), и описанный Коротневым (1888) Gastrodes, тоже гаструлообразный организм, живущий в покровах сальп. Наиболее близкой к гаструле формой являются паразитические организмы, описанные многими исследователями, но далеко не согласно, и именно организмы, живущие в кровеносных сосудах и в полости тела звездчатых червей (Sipunculus, Phymosoma). По описанию Кюнстлера и Грювеля (K ü nstler & Gruvel, 1897), эти организмы являются двуслойными и легко сравнимыми с гаструлой, но другие считают эти формы не за самостоятельные организмы, а за части червей, в которых они найдены (за сложные фагоциты). Если гаструлярная теория находит косвенную опору в этих переходных формах, то и гипотеза Мечникова опирается на два организма, как бы олицетворяющих гипотетическую паренхимеллу. Один из этих организмов — Trichoplax adhaerens — наблюдался Шульце (1883 и 1891) в грацском аквариуме, а другой — Treptoplax reptans (см.) — Монтичелли (1896) в неаполитанском. Оба представляют ползающие пластинки, постоянно меняющие свою форму, и оба являются двуслойными. Эктодерма спинной стороны отлична от таковой брюшной. У Treptoplax она на спинной стороне не имеет ресничек, а на брюшной имеет. У Trichoplax она вся мерцательная, но клетки ее плоские на спинной поверхности и цилиндричны на брюшной. Весь промежуток между этими слоями заполнен похожей на соединительную ткань паренхимой с различными включениями. Оба организма могут размножаться делением. Мечников считает эти организмы за типичную паренхимеллу. Но по некоторым указаниям — под эктодермой Trichoplax наблюдаются мышечные волокна, и даже есть указания (Noll, 1890) на присутствие слухового пузырька. Эти данные, если бы они были достоверны, показывали бы, что мы имеем дело, вероятно, с гораздо более высокой формой вроде бескишечных турбеллярий из низших червей, но эти данные не только не были подтверждены, а отрицаются другими наблюдателями. Существуют еще гипотезы Бючли (Placula-theorie) и Гётте-Заленского (генитогаструлярная), но мы остановимся еще только на гипотезе, получившей разработку в последнее время благодаря открытию загадочного организма — Salinella salve. Этот организм является свободным, но живет в концентрированных соляных растворах, след., тоже в ненормальных условиях. Френцель наблюдал его в Аргентине в сосудах с соленой грязью. Организм представляет однослойный мешок с двумя отверстиями, ротовым и заднепроходным. Можно отличить брюшную и спинную стороны: последняя имеет реснички более редкие. Стенка тела состоит из крупных, кубических клеток, несущих мерцательные волоски как на наружной, так и на внутренней поверхности. Таким образом, этот организм представляет как бы двусимметричную бластулу, но получившую ротовое и заднепроходное отверстия. Самое замечательное, если точны данные Френцеля, размножение этого организма: две особи сходятся, окружаются цистой; клетки их, как предполагает Френцель, вероятно, копулируют между собой по две, а потом каждая клетка по выходе из цисты дает одноклеточный организм, чрезвычайно похожий по форме на взрослую многоклеточную Salinella. Дальнейшее развитие неизвестно. Если сравнить одноклеточного зародыша Salinella со взрослой многоклеточной формой, то сходство в организации действительно поразительно. Седжвик поэтому повторил высказанное еще в 1877 г. Иерингом предположение, что Metazoa могли произойти не последовательным делением одной клетки. а дифференцировкой одной клетки, распадающейся сразу на многоклеточный комлпекс. Стоит допустить, что одноклеточный зародыш Salinella, сохраняя свою форму и вид, стал многоядерным, а потом и много клеточным, и мы получим взрослую Salinella. Однако мы даже не знаем, насколько можно считать описанный Френцелем зародыш за нормальный. Опыты показывают, что в концентрированной морской воде как деление клетки, так и ядра может быть задержано. Весьма возможно, что зародыш Salinella представляет собой именно такое ненормальное явление задержки в делении как ядра, так и самой клетки, составляющей его тело. Таким образом, хотя вопрос о происхождении Metazoa покуда все-таки остается далеким от окончательного и точного решения, но все же теоретически мы можем представить себе происхождение их из колонии простейших. Прочие Mesozoa, как Dicyemidae и Orthonectidae, являются, вероятно, формами, упростившимися под влиянием паразитизма. Чрезвычайно разнообразные мнения были высказаны о происхождении губок. Ввиду значительных затруднений, возникающих при сравнении губок с кишечнополостными, некоторыми (напр. Minchin, 1899) высказывается взгляд, что между пластами губок и прочих Metazoa нет никакой гомологии и что губки могли произойти самостоятельно от простейших, представляющих, подобно Рrоtospongia, колонию воротничковых клеток, как это предполагали и ранее (Carter, 1878; S. Kent, 1881). Во всяком случае губки составляют особую ветвь Metazoa. Переход от кишечнополостных к червям может быть прослежен довольно последовательно. Высшие кишечнополостные от многолучевой симметрии переходят к двулучевой, и стоит себе представить, что ротовое отверстие и органы чувств постепенно перемещаются на тот конец, который при движении (ползании) будет передним, что обособляется брюшная и спинная сторона, и мы получим червеобразный организм. На этой почве и построена гипотеза Ланга о происхождении низших червей от гребневиков (см. Radiata). Переходными формами служат найденная Ковалевским в Красном море Coeloplana metshnikowii и Коротневым, ок. Зондских о-вов, Ctenoplana kowalevskii. Первая из них представляет скорее ползающую турбеллярию с находящимся на брюхе ротовым отверстием и непрерывным мерцательным покровом. Тело ее сплюснуто, подобно телу плоских червей, но у нее сохранились, как у гребневиков, два втяжных щупальца. Рот ведет в 4-лопастное расширение, от которого отходят многочисленные образующие между собой сеть ветви. На аборальной стороне есть отоцист, окруженный венчиком мелких щупалец. Ctenoplana тоже ползающая форма, но сохранила, кроме щупалец, еще 8 мерцательных гребней, могущих втягиваться в желобковидные углубления кожи. Уиллей нашел половозрелую форму Ctenoplana (с мужскими продуктами), и в отличие от Ctenophora, y которых половые продукты падают в гастроваскулярные каналы, у Ctenoplana они выходят через несколько отверстий, лежащих на спинной стороне, наружу. Вероятно, Ctenoplana, как и Ctenophora, гермафродитна. Другая гипотеза происхождения двусимметричных форм от радиальных принадлежит Седжвику (Sedgwik, 1884) и производит двусимметричные членистые формы от сцифополипов. У них ротовое отверстие действительно часто вытягивается по одной оси, около которой радиальные перегородки располагаются симметрично, справа и слева, а равно двулучевая симметричность выражается и в расположении мышц на перегородках. Седжвик допускает, что вытянутый в виде щели рот сцифополипа зарос в средней своей части, но остались два отверстия на краях щели: переднее — ротовое, и заднее — анальное (см. Рот). Если предположить, что расположенные около рта кольцом нервные клетки образовали вследствие описанного процесса два ствола и эта сторона стала брюшной, а на переднем конце образовалось впереди рта скопление нервного вещества, то мы получим схему нервной системы двусимметричных форм. Пространства, лежащие между перегородками, превратились в два ряда полостей, лежащих по бокам тела и представляющих собой целом. Щупальца могли видоизмениться в конечности. Главное неудобство этой гипотезы то, что она связывает членистых вторичнополостных червей непосредственно с радиальными формами и не объясняет нам происхождения низших первичнополостных и бесполостных червей. Можно с большой дозой вероятности предположить, что тип червей был исходным для возникновения всех прочих типов, причем некоторыми придается особое значение личиночной форме червей Trochophora (см.) и гипотетические сходные с ней формы (Trochozoon) считают предками некоторых типов. Однако против этой гипотезы было высказано немало возражений: что касается до иглокожих, то их пятилучевая симметрия есть позднейшее приобретение. Голотурии между ними являются червеобразными формами, и их оральная сторона в то же время передняя, а иногда можно у них отличить брюшную и спинную стороны (по иному расположению амбулакральных ножек на спинной стороне). Семон (1888) полагает, что голотурии суть формы наиболее первичные, и давно их сближает с некоторыми вторичнополостными червями, но многие принимают, наоборот, что голотурии являются формами позднейшими, а первичными считают некоторые ископаемые родственные морским лилиям формы. Возможно, однако, рассматривать амбулакральный зачаток (часто бывающий двойным) как передний целомический сегмент, и тогда предков иглокожих надо искать между вторичнополостными червями с малым числом сегментов, вроде Enteropneusta, или между олигомерными червями. Что касается до моллюсков, то хотя личинка большинства их чрезвычайно напоминает трохофору червей и отличается лишь второстепенными признаками, но весьма вероятно, что моллюски также произошли, по-видимому, от каких-то червеобразных вторичнополостных, но притом, вероятно, нечленистых форм, но указать — от каких именно, мы покуда не можем. Впрочем, некоторые (Pelseneer, 1899) принимают, что предками моллюсков могли быть и членистые формы. В присутствии у некоторых моллюсков (головоногих, хитонов) двух пар метанефридиев видят следы метамерии; с другой стороны, возможно допустить, что вторая пара метанефридиев произошла удвоением первой. Если же признавать филогенетическое значение за трохофорой, то возможно допущение, что моллюски обособились от первичнополостных, подобных трохофоре форм. Наконец, многие принимают, что исходной формой для моллюсков надо считать плоских бесполостных червей, с которыми моллюски представляют некоторое сходство по строению нервной системы, ибо некоторым плоским червям свойственны также 4 нервных ствола, как и моллюскам сем. Chitonidae, a именно пара брюшных и пара боковых стволов (L a ng, 1900). Первое из этих предположений представляется все-таки более вероятным. Вопрос о происхождении членистоногих должен быть расчленен. Происхождение трахейных (см.) s. str., т. е. многоножек и насекомых, от многочленистых, или полимерных, червей стоит вне сомнения благодаря присутствию в современной фауне переходных форм, а именно первичнотрахейных (см.). Сложнее вопрос о происхождении ракообразных и паукообразных. При решении этого вопроса придают большое значение личинке низших ракообразных — Naupl i us, которую иногда сравнивают с трохофорой. Сравнение Nauplius с трохофорой затруднительно прежде всего потому, что первая из этих личинок является членистой, а вторая в ее типичной и первоначальной форме — нечленистой. Так как мы не можем определить с точностью, сколько пар конечностей исчезло у ракообразных из числа лежащих впереди рта (см. Усики), то мы и не можем сказать, из скольких сегментов состоит Nauplius, но во всяком случае число это более числа наличных конечностей его, т. е. более трех. Таким образом, если искать на основании этих данных сравнения между червями, то можно было бы сравнивать его с вторичнополостными, но с небольшим числом члеников червями вроде Dinophilus. У этого последнего отношение полости тела к метанефридиям действительно напоминает таковое членистоногих. Точно так же сходство между Dinophilus и низшими ракообразными выражается в одновременном присутствии у него и у некоторых из ракообразных как вторичной, так и первичной полости тела, причем последняя не приняла еще характера кровеносной системы. Сходство выражается в присутствии у Dinophilus зачатка мезоскелетной пластинки, соответствующей поперечному мускулу, смыкающему у ракообразных створки панциря. Одним словом, весьма вероятно, что предками ракообразных являются вторичнополостные черви с небольшим числом сегментов и что Nauplius представляет собой повторение такой динофилоподобной стадии. Червей таких можно назвать мезомерными в отличие от олигомерных и полимерных. Потом число члеников у ракообразных возросло, но эти членики по способу своего образования и онтогенического развития весьма сильно разнятся от члеников науплиуса, что и указывает на их позднейшее филогенетическое происхождение. Весьма вероятно, что мезомерные черви были родоначальниками не только ракообразных, но и таких загадочных форм, как Tardigrada, имеющих при небольшом числе сегментов ножки с рудиментарной членистостью, а может быть, и Pantopoda, представляющих собой уже типичных членистоногих. Происхождение паукообразных ставят в связи с существованием в современной и ископаемой фауне особой группы членистоногих — Меrostomata, которую возможно связать с низшими ракообразными. Многочисленные гипотезы имеются касательно происхождения хордовых, но из них остановимся на двух. Одна из них (Семпер и Дорн) предками хордовых считает полимерных червей, причем принимает, что спинная сторона хордовых, где лежит нервная трубка, соответствует брюшной стороне червей, на которой у них тоже лежит нервная цепочка. Другая гипотеза (Бетсон, Шимкевич и др.) производит хордовых от олигомерных червей, как кишечножаберные (см.), имеющих жаберные щели, короткую спинную нервную трубку, а по мнению некоторых, и хорду, т. е. ряд признаков, сближающих их с хордовыми (см.). Таким образом, Ф. типов и крупных групп животного царства в грубой схеме может быть представлена следующим образом; конечно, эта схема весьма гипотетична и до известной степени субъективна.
Литература приведена в "Биологических основах зоологии" Шимкевича (СПб., 1901).
В. Шимкевич.